Domain swapping reveals that the N-terminal domain of the sensor kinase KdpD in Escherichia coli is important for signaling
09-Jul-2009
BMC Microbiology, 2009, 9(133), doi:10.1186/1471-2180-9-133 published on 09.07.2009
BMC Microbiology, online article
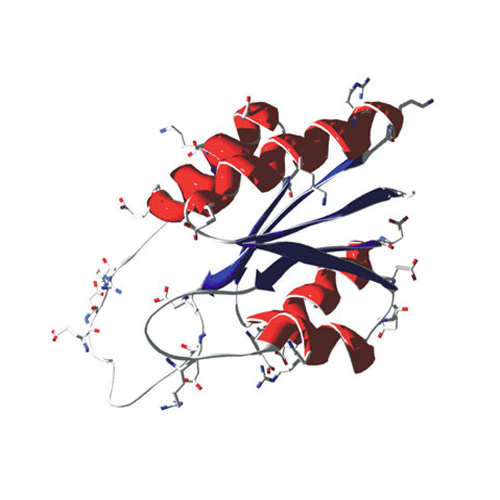
The KdpD/KdpE two-component system of Escherichia coli regulates expression of the kdpFABC operon encoding the high affinity K+ transport system KdpFABC. The input domain of KdpD comprises a domain that belongs to the family of universal stress proteins (Usp). It has been previously demonstrated that UspC binds to this domain, resulting in KdpD/KdpE scaffolding under salt stress. However the mechanistic significance of this domain for signaling remains unclear. Here, we employed a "domain swapping" approach to replace the KdpD-Usp domain with four homologous domains or with the six soluble Usp proteins of E. coli. Full response to salt stress was only achieved with a chimera that contains UspC, probably due to unaffected scaffolding of the KdpD/KdpE signaling cascade by soluble UspC. Unexpectedly, chimeras containing either UspF or UspG not only prevented kdpFABC expression under salt stress but also under K+ limiting conditions, although these hybrid proteins exhibited kinase and phosphotransferase activities in vitro. These are the first KdpD derivatives that do not respond to K+ limitation due to alterations in the N-terminal domain. Analysis of the KdpD-Usp tertiary structure revealed that this domain has a net positively charged surface, while UspF and UspG are characterized by net negative surface charges. The Usp domain within KdpD not only functions as a binding surface for the scaffold UspC, but it is also important for KdpD signaling. We propose that KdpD sensing/signaling involves alterations of electrostatic interactions between the large N- and C-terminal cytoplasmic domains.